Межвидовая гибридизация иногда приводит к появлению новых видов не только у растений, но и у животных. Генетические основы этого процесса изучены слабо. Анализ геномов южноамериканских бабочек показал, что вид Heliconius elevatus возник 180 тысяч лет назад в результате гибридизации H. pardalinus и H. melpomene. Гибридный вид унаследовал от первого родительского вида 99% генома, а от второго лишь 1%, однако в этом одном проценте сконцентрированы важные гены, связанные с экологической специализацией, окраской крыльев, выбором брачного партнера и репродуктивной изоляцией. Гибридный вид продолжает обмениваться генами с H. pardalinus, но не сливается с ним, по-видимому, благодаря отбору, который отсеивает особей с неполным комплектом признаков, унаследованных от H. melpomene. Ранее предполагалось, что отбор может справиться с такой задачей, только если ключевые гены собраны в небольшое число кластеров («островков видообразования») и защищены от рекомбинации инверсиями. Но у H. elevatus островков видообразования много, а инверсий нет. Исследование показало, что возможные сценарии симпатрического видообразования более разнообразны, чем считалось.
Центрально- и южноамериканские бабочки рода Heliconius — популярный объект эволюционных исследований (см. ссылки в конце новости). Эволюция узоров на их крыльях — классический пример мюллеровской мимикрии. Бабочки Heliconius несъедобны из-за цианогенных гликозидов, которые отчасти происходят из растений семейства Passifloraceae, которыми питаются гусеницы, а отчасти синтезируются самими бабочками. У этой защиты, однако, есть один недостаток, который как раз и способствует эволюции мюллеровской мимикрии. Птицы от рождения не знают, какие виды разноцветных бабочек съедобны, а какие нет. Они выясняют это в течение жизни на собственном горьком опыте.
Но пока птицы учатся, бабочки погибают. Поэтому отбор поддерживает у несъедобных бабочек такие вариации окраски, которые делают их похожими на других несъедобных бабочек, многочисленных в данном районе. Это повышает шансы, что птица не тронет бабочку, поскольку уже «обожглась» на похожей добыче.В результате разнообразие узоров на крыльях бабочек распадается на кластеры, поддерживаемые дизруптивным отбором. Особи, принадлежащие к тому или иному кластеру, оказываются в выигрыше, а промежуточные варианты проигрывают, потому что птицы их не узнают.
Окрасочные кластеры (типы узоров) соответствуют изолированным пикам на ландшафте приспособленности (Fitness landscape). При этом не очень понятно, как бабочки переходят с одного пика на другой. Одним из путей теоретически может быть гибридогенное видообразование.
У животных межвидовая гибридизация гораздо реже приводит к появлению новых видов, чем у растений (возможно, потому, что растения легче переносят полиплоидизацию, которая радикально облегчает процесс гибридогенного видообразования, но это тема для отдельного разговора). Но все же иногда такое случается и у животных, причем без всякой полиплоидизации (см. Новый вид дарвиновых вьюрков возник на глазах у исследователей, «Элементы», 04.12.2017).
О генетических механизмах и сценариях гибридогенного видообразования у животных известно немного. Интересные новые данные об этом получил коллектив биологов из США, Великобритании, Перу, Швеции, Венесуэлы, Франции, Панамы, Бразилии, Канады, Колумбии и Германии в ходе изучения геномов трех южноамериканских видов Heliconius: H. elevatus, H. melpomene и H. pardalinus.
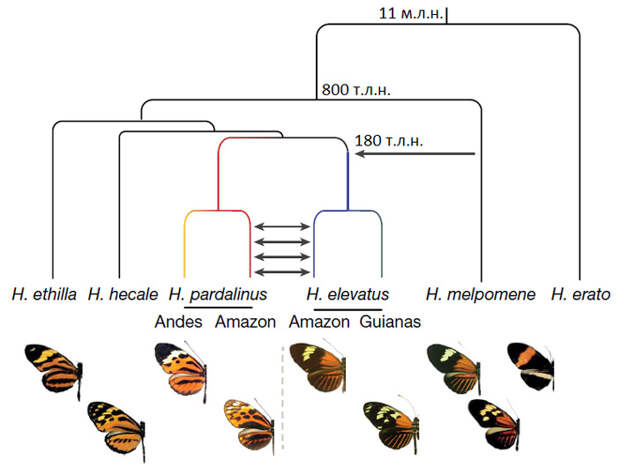
Эти виды привлекли внимание исследователей, потому что ранее было показано, что H. elevatus генетически близок к H. pardalinus, хотя по окраске он похож на гораздо более далекий вид H. melpomene. У H. pardalinus и нескольких его близких родственников тип окраски «тигровый», а у H. elevatus и H. melpomene — «красно-черно-желтый» (рис. 1). При этом все три вида являются симпатрическими (проживают совместно) на огромной территории Амазонии (рис. 2). Эти факты позволили предположить, что вид H. elevatus возник в результате межвидовой гибридизации.
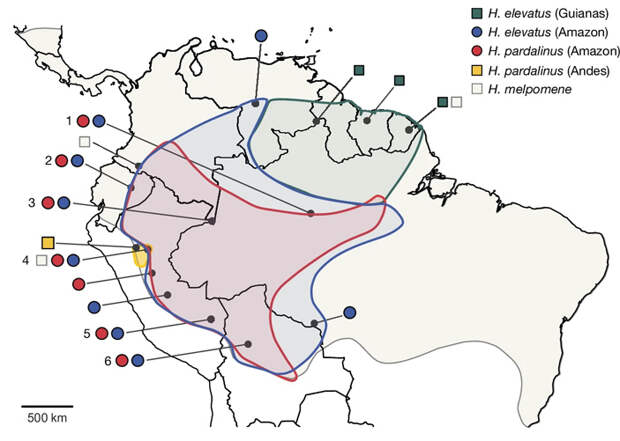
Чтобы проверить эту гипотезу, авторы отсеквенировали полные геномы 92 бабочек (42 особи H. elevatus, 33 H. pardalinus и 17 H. melpomene), пойманных как в симпатрических, так и в аллопатрических частях их ареалов (рис. 2). К этим геномам применили всю мощь современных методов популяционной генетики и сравнительной геномики — так же, как это делают с человеческими геномами, когда реконструируют историю человеческих популяций и выясняют, где, когда и с кем скрещивались их предки.
Один из подходов состоял в том, что строились филогенетические деревья по отдельности для каждого из 11 509 непересекающихся кусочков генома, в каждом из которых находится по 1000 снипов (однонуклеотидных полиморфизмов). В большинстве случаев на получившихся деревьях вид H. elevatus группируется с H. pardalinus, а H. melpomene стоит особняком. Однако на некоторых деревьях H. elevatus оказывается ближе к H. melpomene. Это согласуется с гибридным происхождением H. elevatus. При этом на деревьях, демонстрирующих родство H. elevatus и H. pardalinus, симпатрические популяции этих видов, как правило, оказываются ближе друг к другу, чем представители одного и того же вида из разных частей ареала. Это указывает на продолжающийся обмен генами между H. elevatus и H. pardalinus в районах совместного проживания. С другой стороны, по тем участкам генома H. elevatus, по которым он сходен с H. melpomene, бабочки почти всегда группируются с конспецификами из других районов, а не с представителями другого вида из того же района. Это значит, что обмена генами между H. elevatus и H. melpomene в настоящее время не происходит.
На основе этого и множества других тестов ученые пришли к выводу, что вид H. elevatus появился около 180 тысяч лет назад в результате гибридизации H. pardalinus и H. melpomene. Большую часть наследственного материала гибридный вид получил от H. pardalinus, а от H. melpomene ему досталось лишь около 1% генома. Гибридный вид репродуктивно изолирован от H. melpomene, но с другим родительским видом, H. pardalinus, он продолжает иногда скрещиваться. Гибридизация происходит в Амазонии, где все три вида являются симпатрическими. Речь может идти о нескольких случаях плодовитых гетероспецифических скрещиваний в каждом поколении, или даже о нескольких десятках. Это многовато для сохранения генетической обособленности, даже несмотря на то, что эффективная численность амазонских популяций H. elevatus и H. pardalinus исчисляется миллионами.
Продолжающаяся гибридизация, однако, не приводит к слиянию H. elevatus и H. pardalinus. Эти виды четко различаются и по предпочитаемым кормовым растениям, и по окраске и форме крыльев, и по брачным предпочтениям (самцы предпочитают ухаживать за «своими» самками не только в природе, но и в экспериментах с искусственными самками, различающимися только по окраске крыльев), и даже по динамике полета (H. elevatus машут крыльями чаще: в среднем 11,3 взмахов в секунду против 10,8 у H. pardalinus).
Как же удается двум видам сохранять морфологическую и экологическую обособленность, невзирая на постоянный обмен генами?
Нужно пояснить, что это далеко не единственный случай, когда пары симпатрических видов сохраняют свою идентичность при неполной репродуктивной изоляции (C. Pinho, J. Hey, 2010. Divergence with Gene Flow: Models and Data). В тех редких случаях, когда генетику таких пар удается изучить достаточно подробно, обычно обнаруживается небольшое число участков генома с резко пониженным сходством. Их называют «геномными островками дивергенции» или «островками видообразования» (см. ссылки в конце новости). Если ключевые гены, ответственные за межвидовые различия, собраны в небольшое число компактных кластеров, то соответствующие фенотипические признаки не так сильно перемешиваются при гибридизации, и отбору легче справляться с отбраковкой особей со смешанной морфологией и экологией. Иногда «островки видообразования» дополнительно защищены от рекомбинации инверсиями (см. Межвидовая гибридизация способствует накоплению хромосомных инверсий, «Элементы», 04.09.2017).
Отталкиваясь от этих фактов, исследователи попытались найти островки видообразования в геномах H. elevatus и H. pardalinus. Для каждого участка генома вычислили показатель FST (см. Fixation index; это стандартный способ количественной оценки дивергенции популяций). Оказалось, что для 99% генома значение FST в данной паре видов близко к нулю, как если бы они были единой популяцией. Области с повышенным FST составляют менее 1% генома и образуют 44 «островка дивергенции», разбросанных почти по всем хромосомам. Обнаружение островков дивергенции было ожидаемым результатом, а столь большое их число — неожиданным. Обычно островков бывает меньше. Еще одним неожиданным результатом оказалось отсутствие инверсий, которые у других пар недавно разделившихся симпатрических видов часто защищают островки дивергенции от рекомбинации и распада.
Также выяснилось, что в пределах островков дивергенции резко повышена доля участков, унаследованных от H. melpomene. Такие участки обнаружены в 32 островках из 44, что гораздо больше ожидаемого при случайном распределении. Таким образом, гибридный вид H. elevatus, по-видимому, получил от H. melpomene некие ключевые гены, позволяющие ему сохранять свою видовую идентичность и не сливаться с H. pardalinus, несмотря на несовершенство репродуктивных барьеров.
Чтобы проверить это предположение, ученым пришлось провести колоссальную работу по картированию признаков, различающих изучаемые виды. Для этого представителей симпатрических амазонских популяций H. elevatus и H. pardalinus скрещивали в лаборатории. Обычно они не желают скрещиваться, но если выбора нет, то все-таки скрещиваются, и потомство получается плодовитое. Во втором поколении гибридов, как положено по законам генетики, исследователи получили множество разнообразных комбинаций видовых признаков. Этих бабочек генотипировали и стали искать аллели, от которых зависит, на какой из двух родительских видов похож гибридный потомок по данному признаку. В итоге удалось выявить 63 локуса (QTL, см. Локус количественных признаков), аллельное состояние которых объясняет значительную часть изменчивости гибридного потомства по признакам, отвечающим за видовую идентичность и репродуктивную изоляцию. Среди этих признаков — окраска и форма крыльев, набор мужских половых феромонов, частота махания крылышками (этот признак тоже учитывается бабочками при выборе партнера), мужские предпочтения при выборе самок по окраске крыльев и женские предпочтения при выборе кормового растения. По большинству из этих признаков гибридный вид H. elevatus больше похож на H. melpomene, с которым он не скрещивается, чем на H. pardalinus, с которым он нехотя, но скрещивается.
В итоге оказалось, что гены, отвечающие за эти признаки, как и ожидалось, с повышенной частотой обнаруживаются в участках генома H. elevatus, унаследованных от H. melpomene.
Таким образом, вырисовывается следующая картина. Вид H. elevatus возник 180 тысяч лет назад в результате редкого события — гибридизации двух довольно далеких (разошедшихся 800 тысяч лет назад) видов H. melpomene и H. pardalinus. Гибридное потомство получило такую комбинацию морфологических и экологических признаков, которая сразу поместила их на отдельный пик ландшафта приспособленности (авторы, к сожалению, не сообщают, чем конкретно H. elevatus отличается от H. melpomene по экологии, но сильные генетические различия в 99% генома позволяют предполагать, что различий достаточно). Эта комбинация обеспечила надежную репродуктивную изоляцию от H. melpomene (опять-таки, авторы не исследовали механизмы этой изоляции, удовлетворившись тем фактом, что в геномах нет признаков продолжающегося обмена генами между H. elevatus и H. melpomene, несмотря на симпатрию).
От другого родительского вида, H. pardalinus, гибридный вид H. elevatus отделен несовершенным (то есть проницаемым) репродуктивным барьером. Из-за этого 99% генома H. elevatus выглядят так, как будто принадлежат единому генофонду с H. pardalinus. По сути, так оно и есть: постоянные межвидовые скрещивания перемешивают эту часть генома двух видов почти до полной гомогенности.
Однако H. elevatus сохраняет свою видовую идентичность и экологическую обособленность от H. pardalinus за счет оставшегося одного процента генома. Этот один процент разбросан по разным хромосомам в виде 44 кластеров (островков дивергенции), которые не участвуют в генетическом обмене с H. pardalinus. В этих кластерах сосредоточены ключевые гены, полученные гибридным видом от H. melpomene. Эти гены влияют на такие важные для видообразования признаки, как окраска крыльев, брачные предпочтения, половые феромоны и выбор кормового растения (бабочки Heliconius спариваются рядом со своим кормовым растением, поэтому последний признак тоже важен для репродуктивной изоляции).
Ранее предполагалось, что для успешного симпатрического видообразования при сохраняющемся обмене генами необходимо, чтобы ключевые гены были собраны в небольшое число компактных островков дивергенции, желательно еще и защищенных от перемешивания инверсиями. Думали, что иначе отбор не справится с отбраковкой всех возникающих неудачных комбинаций видовых признаков, так что в итоге либо виды сольются, либо один из них вымрет. Но на данном примере мы видим, что симпатрическое видообразование с сохраняющимся потоком генов возможно и с большим числом островков, и без инверсий. Отбор как-то справляется, и виды H. elevatus и H. pardalinus продолжают существовать как самостоятельные виды уже около двухсот тысяч лет несмотря на дырявый репродуктивный барьер. Из этого следует, что симпатрическое видообразование в природе может происходить чаще, чем предсказывают имеющиеся модели.
Источник: Neil Rosser, Fernando Seixas, Lucie M. Queste, Bruna Cama, Ronald Mori-Pezo, Dmytro Kryvokhyzha, Michaela Nelson, Rachel Waite-Hudson, Matt Goringe, Mauro Costa, Marianne Elias, Clarisse Mendes leres de Figueiredo, André Victor Lucci Freitas, Mathieu Joron, Krzysztof Kozak, Gerardo Lamas, Ananda R. P. Martins, W. Owen McMillan, Jonathan Ready, Nicol Rueda-Muñoz, Camilo Salazar, Patricio Salazar, Stefan Schulz, Leila T. Shirai, Karina L. Silva-Brandão, James Mallet & Kanchon K. Dasmahapatra. Hybrid speciation driven by multilocus introgression of ecological traits // Nature. 2024. DOI: 10.1038/s41586-024-07263-w.
См. также об эволюции бабочек рода Heliconius:
1) Мимикрирующие друг под друга бабочки-нимфалиды научились при выборе партнера использовать запах, «Элементы», 13.01.2016.
2) Зафиксирован начальный этап видообразования у тропических бабочек, «Элементы», 09.11.2009.
3) Найден ген, отвечающий за эволюцию окраски у бабочек, «Элементы», 31.08.2011.
О «геномных островках видообразования»:
1) Различия между зарождающимися видами африканских цихлид записаны в «геномных островках видообразования», «Элементы», 21.12.2015.
2) Видообразование у коралловых рыбок Hypoplectrus основано на четырех генах, влияющих на зрение и окраску, «Элементы», 11.03.2019.
3) Генетические различия между серой и черной вороной записаны на «островках видообразования», «Элементы», 25.06.2014.
Свежие комментарии