Ключевым событием в эволюции многоклеточности является изменение уровня, на котором работает естественный отбор. Он должен перейти с уровня клеток на уровень групп клеток. При этом группы должны обладать наследственной изменчивостью по признакам, не сводимым к простой сумме свойств составляющих группу клеток. В результате группа становится «индивидом» в эволюционном смысле и получает возможность приобретать собственные адаптации. Американские биологи воспроизвели этот важнейший эволюционный переход в эксперименте на дрожжах. За 600 дней искусственного отбора на скорость оседания на дно пробирки дрожжи научились образовывать макроскопические (различимые невооруженным глазом) агрегаты, состоящие из плотно сплетенных многоклеточных нитей и размножающиеся бесполым путем. Помимо исходной мутации, препятствующей разделению почкующихся клеток, у дрожжей в ходе эксперимента закрепились десятки мутаций, повышающих механическую прочность «организма» и помогающих ему вырасти большим. Это эмерджентные признаки, закодированные в геноме клетки, но проявляющиеся на уровне организма (например, механическую прочность многоклеточного агрегата нельзя измерить у отдельной клетки). Интересно, что столь легкое превращение в крупный организм дрожжи демонстрируют только при условии строгой анаэробности. Доступность аэробного метаболизма приводит к отбору на измельчание, потому что в маленьких агрегатах клетки лучше снабжаются кислородом и быстрее растут. В итоге многоклеточные дрожжи остаются микроскопическими. Результаты эксперимента помогают понять причины миллиардолетней паузы между появлением эукариот и расцветом макроскопических многоклеточных организмов (так называемый «Скучный миллиард»).
Коллектив американских биологов под руководством Уильяма Рэтклиффа (William C. Ratcliff) сообщил о новых результатах опытов с многоклеточными дрожжами-снежинками (snowflake yeast), которыми Рэтклифф с коллегами занимаются уже более десяти лет.
О начальных этапах этих исследований рассказано в новости Экспериментально показано образование многоклеточных эукариот из одноклеточных предков («Элементы», 25.01.2012). Дрожжи-снежинки были выведены путем искусственного отбора на размер клеточных скоплений. Фактически это отбор на скорость оседания. Дрожжи в течение суток (пять-шесть клеточных поколений) живут в жидкой среде при постоянном потряхивании. Потом их оставляют в покое на несколько минут, чтобы они начали оседать на дно. Чем крупнее клеточный агрегат, тем быстрее он оседает. Самая нижняя часть взвеси переносится в пробирку со свежей питательной средой, и цикл повторяется.
У дрожжей-снежинок, выведенных таким способом, клетки не отделяются друг от друга при почковании. На почках вырастают новые почки, и получаются многоклеточные агрегаты диаметром около 0,05 мм, действительно напоминающие снежинки. Они размножаются бесполым путем, отделяя дочерние снежинки. Агрегаты эти очень непрочные, потому что все потомки любой клетки прикрепляются к колонии только через эту клетку — свою прародительницу. Стоит разорваться любой межклеточной связи, и от колонии отделяется вся веточка потомков оторвавшейся клетки. Собственно, именно так эти колонии и размножаются.
Рэтклифф с коллегами изучили мутации, отличающие дрожжи-снежинки от обычных дрожжей. Как выяснилось, для того, чтобы клетки перестали расходится после почкования, достаточно заблокировать работу одного-единственного гена, ACE2 (W. Ratcliff et al., 2015. Origins of multicellular evolvability in snowflake yeast). Это очень удобно, и теперь для моделирования эволюции многоклеточности ученые используют дрожжи-снежинки, полученные из обычных дрожжей (с тем или иным известным исследователю генотипом) путем удаления гена ACE2.
В 2021 году были опубликованы результаты 145-дневного эволюционного эксперимента, в котором дрожжи-снежинки опять отбирались на крупный размер по описанной выше схеме (G. Bozdag et al., 2021. Oxygen suppression of macroscopic multicellularity). В этом исследовании проверялось влияние кислорода и аэробного метаболизма на эволюцию крупных многоклеточных агрегатов. Использовались три разновидности дрожжей-снежинок: строго анаэробные (с мутацией в митохондриальной ДНК, делающей кислородное дыхание невозможным), обычные миксотрофные (способные и к анаэробному сбраживанию глюкозы, и к кислородному дыханию) и строго аэробные (генетически это были такие же дрожжи, как в предыдущем случае, но их кормили не глюкозой, а глицеролом, из которого дрожжи могут извлекать энергию только аэробным путем).
Оказалось, что дрожжи-снежинки охотно эволюционируют в сторону увеличения размера только при строгой анаэробности. Возможность аэробного метаболизма создает новый вектор отбора, направленный в сторону, противоположную искусственному отбору на скорость оседания. В крупных клеточных агрегатах у большинства клеток начинаются проблемы с доступностью кислорода, что приводит к отбору на измельчание колоний. Выигрыш в скорости роста, даваемый кислородным дыханием, настолько велик, что уже при среднем радиусе колоний около 30 мкм он сводит на нет выигрыш от крупного размера, создаваемый искусственным отбором на скорость оседания. Между двумя векторами отбора устанавливается равновесие, и колонии остаются микроскопическими (рис. 2).

Миксотрофные снежинки после 145 раундов отбора стали чуть крупнее (радиус около 40 мкм), анаэробные — еще крупнее (50 мкм). Но самыми крупными (средний радиус колонии 60–70 мкм) выросли аэробные снежинки, которым давали дополнительный кислород. По мнению авторов, эти результаты помогают понять, почему в истории жизни на Земле был миллиардолетний период эволюционного застоя между появлением эукариот (в начале протерозойского эона) и началом расцвета крупных многоклеточных форм жизни (в конце протерозоя). В этот период, известный как «Скучный миллиард», концентрация кислорода в атмосфере и гидросфере оставалась низкой. Возможно, это накладывало жесткие ограничения на эволюцию примитивных многоклеточных эукариот. Лишь ближе к концу протерозоя, когда концентрация кислорода снова стала расти, многоклеточные эукариоты начали бурно эволюционировать и становиться крупнее (см.: Размер живых существ увеличивался скачками, «Элементы», 30.12.2008).
Так или иначе, в 145-дневном эксперименте все дрожжи-снежинки остались микроскопическими, то есть неразличимыми (или почти неразличимыми) невооруженным глазом. В дальнейшем рубеж между микро- и макроскопическим размером удалось преодолеть. Рэтклифф и его коллеги сообщили об этом в новой статье, опубликованной в мае 2023 года в журнале Nature. Авторы намерены продолжать свой эксперимент по отбору дрожжей-снежинок на крупный размер в течение многих лет, вдохновляясь примером Ричарда Ленски. Ну а пока они отчитались о первом 600-дневном этапе.
В эксперименте участвовали те же три варианта культур дрожжей-снежинок, о которых сказано выше: строго аэробные, миксотрофные и анаэробные. Каждый вариант представлен пятью линиями (повторностями), то есть в общей сложности было 15 независимо эволюционирующих линий. Опыты с подачей дополнительного кислорода в новой статье не упоминаются.
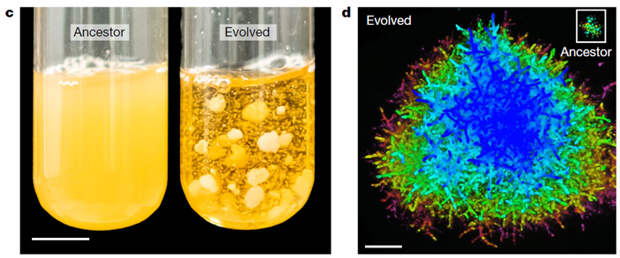
Аэробные и миксотрофные линии, не достигшие макроскопических размеров за 145 дней отбора, не сумели этого сделать и за 600 дней. А вот анаэробные — сумели, причем во всех пяти повторностях. Средний радиус колоний увеличился до 450 мкм, а в одной из повторностей — почти до 600 мкм. Отдельные колонии достигают нескольких миллиметров в диаметре, то есть они крупнее, чем мушка-дрозофила, и, конечно, они прекрасно видимы невооруженным глазом (рис. 1). Среднее число клеток в кластере увеличилось от примерно 100 у предковых дрожжей-снежинок до полумиллиона.
Большая часть статьи посвящена поискам ответа на вопрос, как именно — за счет каких морфологических, биофизических и генетических изменений — анаэробным дрожжам-снежинкам удалось стать такими крупными.
Как выяснилось, ключевые изменения затронули, во-первых, форму клеток: из округлых они стали сильно вытянутыми. Во-вторых, основание почки (bud neck) стало намного шире, что повысило прочность соединения почки с родительской клеткой. В-третьих, у многоклеточных агрегатов появилась принципиально новая особенность: цепочки клеток, ставшие похожими на гифы нормальных многоклеточных грибов, начали сплетаться друг с другом.
Изначально округлые клетки становились всё более и более вытянутыми на протяжении всего эксперимента (рис. 3). Это имеет прямое отношение к увеличению размера колонии. Дело в том, что удлиненные клетки в ходе последовательного почкования легче упаковываются в пространстве, не расталкивают друг друга и не создают избыточного напряжения, ведущего к разрыву межклеточных связей.
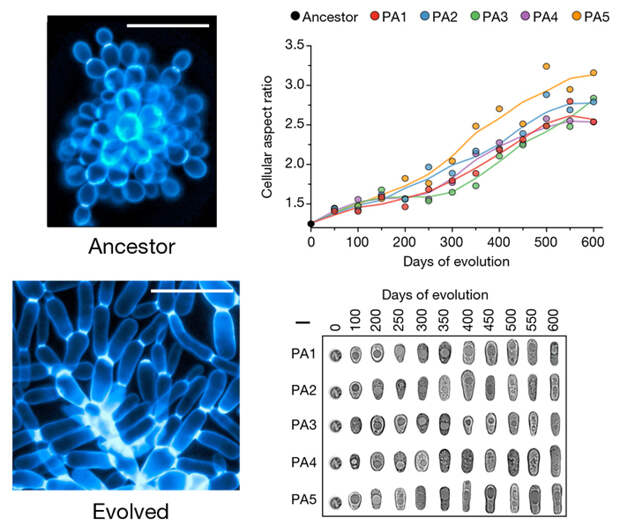
Начиная с определенного момента, когда отношение длины клетки к ее ширине приближается к двум (этот порог во всех пяти повторностях был достигнут между 300-м и 400-м днями эксперимента), количественные изменения переходят в качественные: ветвящиеся многоклеточные «гифы», из которых состоит колония, начинают сплетаться друг с другом.
Это резко повышает плотность упаковки клеток, а главное — механическую прочность колонии. Теперь колония уже не распадается от разрыва любой межклеточной связи. Как и у предковых дрожжей-снежинок, такой разрыв ведет к образованию топологически отдельной веточки. Но только теперь она с большой вероятностью останется присоединена к колонии благодаря плотному сплетению гиф. Крупные колонии перестают представлять из себя топологически целостные объекты, состоящие из непрерывных цепочек клеток. Они складываются из множества фрагментов, прочно сплетенных друг с другом. Интересно, что при этом сохраняется моноклональная природа колоний, характерная для предковых дрожжей-снежинок. Это значит, что каждый многоклеточный агрегат состоит из потомков одной клетки (мы ведь помним, что отрывающиеся веточки, служащие для размножения колоний, состоят из одной родительской клетки и ее потомков). Почему же гифы разных колоний не сплетаются друг с другом? Ключевую роль в этом играет постоянное перемешивание жидкой культуры. Если оставить пробирку в покое, разнородные колонии прекрасно друг с другом сплетаются, образуя колонии-химеры, что не очень хорошо с точки зрения эволюционных перспектив многоклеточного организма. Моноклональность обеспечивает генетическую идентичность клеток организма и снимает проблему их эгоистической эволюции (см.: Выращивание монокультур — ключ к эффективности сельского хозяйства у термитов, «Элементы», 24.11.2009).
Авторы изучили влияние сплетенности (entanglement) и усиления межклеточных соединений на механические свойства колоний. Оказалось, что по сравнению с предковыми дрожжами-снежинками их прочность выросла на много порядков. В статье приведены следующие цифры. Многоклеточные колонии предков разрушаются при механическом напряжении 240 Па и имеют ударную вязкость 8,9 Дж/м3. У крупных колоний, полученных за 600 дней отбора, соответствующие значения — 7 МПа и 0,6 МДж/м3. В переводе на человеческий язык, прочность предковых колоний — примерно как у желе, а у проэволюционировавших — примерно как у древесины.
Авторы отсеквенировали геномы подопытных дрожжей и выявили 123 мутации, закрепившиеся в пяти анаэробных линиях. В разных линиях закрепились разные мутации, хотя некоторые из них затрагивают одни и те же гены. Общими для всех пяти линий являются функции генов, затронутых мутациями. Многие мутации, как и следовало ожидать, изменили кодирующие или регуляторные области генов, о которых известно, что они влияют на динамику почкования, рост клеток, их форму и ширину основания почки. Таким образом, параллельная эволюция в пяти повторностях пришла к похожим фенотипическим результатам за счет разных генетических изменений. Это логично, учитывая мультигенность признаков, на которые был направлен отбор.
Эксперимент показал, с какой легкостью одноклеточные грибы могут переходить к многоклеточности и приобретать способность к построению прочных макроскопических структур из переплетенных гиф — способность, которая в ходе эволюции грибов возникала много раз независимо. Для этого оказалось достаточно простой экспериментальной манипуляции, сводящейся к тому, чтобы поставить выживание клеток в зависимость от размера групп, в которые эти клетки входят. В природе такой отбор могут осуществлять хищники, поедающие отдельные клетки, но не справляющиеся с крупными клеточными скоплениями. Показано, например, что такие хищники стимулируют эволюцию колониальности у одноклеточных водорослей (M. Boraas et al., 1998. Phagotrophy by a flagellate selects for colonial prey: A possible origin of multicellularity).
В эксперименте отбор на размер моноклональных клеточных групп фактически сразу сделал эти группы «дарвиновскими индивидами», то есть эволюционирующими объектами, вырабатывающими собственные адаптации. В результате такого отбора в геномах клеток послушно закрепляются мутации, помогающие развиваться «эмерджентным» групповым признакам, не сводимым к простой сумме наследственных свойств клеток. Хотя, конечно, эти групповые признаки в ходе онтогенеза колонии развиваются исключительно на основе свойств отдельных клеток. В эксперименте в роли признаков надклеточного (организменного) уровня выступали размер колонии и ее механическая прочность, определяемая шириной основания почки и степенью переплетенности гиф, которая, в свою очередь, во многом определялась формой клеток.
Крайне интересен и подтвердившийся вывод о том, что аэробный метаболизм при ограниченной доступности кислорода может стать серьезным препятствием для развития примитивной многоклеточности. Может быть, этот вывод действительно в какой-то мере объясняет загадку «Скучного миллиарда». Если эксперимент и в самом деле станет долгосрочным, как у Ричарда Ленски, то он наверняка поможет лучше разобраться в механизмах и факторах перехода эукариот к многоклеточности.
Источник: G. Ozan Bozdag, Seyed Alireza Zamani-Dahaj, Thomas C. Day, Penelope C. Kahn, Anthony J. Burnetti, Dung T. Lac, Kai Tong, Peter L. Conlin, Aishwarya H. Balwani, Eva L. Dyer, Peter J. Yunker & William C. Ratcliff. De novo evolution of macroscopic multicellularity // Nature. 2023. DOI: 10.1038/s41586-023-06052-1.
См. также:
Экспериментально показано образование многоклеточных эукариот из одноклеточных предков, «Элементы», 25.01.2012.
Свежие комментарии