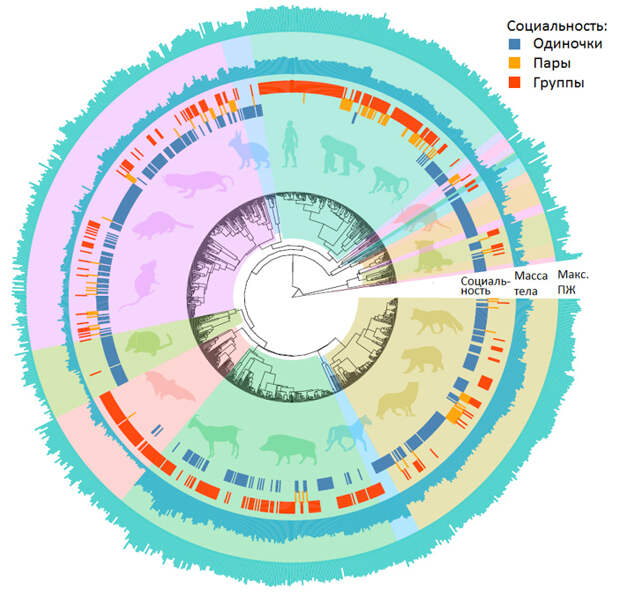
Сравнительный анализ данных по продолжительности жизни и социальному поведению 974 видов млекопитающих подтвердил положительную связь между социальностью и долголетием: со всеми необходимыми поправками получилось, что социальные звери живут дольше одиночек. Эволюционные переходы от короткоживущего состояния к долгоживущему чаще происходят у социальных млекопитающих, чем у одиночек, однако переходы от одиночной жизни к социальности происходят с одинаковой частотой у коротко- и долгоживущих. Это значит, что сопряженная эволюция социальности и долголетия объясняется не тем, что долголетие способствует отбору на социальность, а тем, что социальность способствует отбору на долголетие.
Идея о том, что общественный образ жизни может способствовать эволюции долголетия (замедленного старения), давно обсуждается в научной литературе. Под эту связь подведен неплохой теоретический базис, о котором рассказано в новости Что общего у голых землекопов и «голых обезьян»? («Элементы», 06.03.2017). Суть в том, что формирующаяся в ходе эволюции у того или иного вида животных скорость старения (понимаемого как рост вероятности смерти с возрастом) зависит от того, насколько быстро падает с возрастом сила отбора против мутаций, снижающих жизнеспособность в этом возрасте и позднее. При высоком уровне «базовой», не связанной с возрастом смертности (например, при сильном прессе хищников) шансы животного прожить долгую жизнь будут низкими, даже если животное вовсе не будет стареть. Чем меньше особей доживает до возраста N, тем слабее отбор против вредных мутаций, проявляющихся в этом возрасте или позже. Поэтому классическая эволюционная теория старения предсказывает, что у видов, хорошо защищенных от внешних угроз, в ходе эволюции должна чаще формироваться низкая скорость старения и высокая продолжительность жизни (ПЖ), а у слабо защищенных — быстрое старение и низкая ПЖ.
Об одном из эмпирических подтверждений этой идеи рассказано в новости Панцири и яды способствуют эволюции долголетия у амфибий и рептилий («Элементы», 05.07.2022).Социальность защищает животных от внешних угроз множеством способов (больше глаз, носов и ушей, чтобы вовремя заметить хищника, пониженная вероятность, что съедят именно вас, если вы пасетесь со стадом, коллективная оборона, взаимопомощь, совместная забота о потомстве и т. д.). Поэтому логично предположить, что у общественных животных в ходе эволюции будет чаще развиваться медленное старение и высокая ПЖ, чем у одиночек. Отдельных примеров, подтверждающих такую тенденцию, известно немало. Например, два самых социальных вида млекопитающих — голые землекопы и люди — по совместительству являются и рекордсменами-долгожителями среди нелетающих зверей своего размерного класса (умение летать — великолепная защита, поэтому многие рукокрылые живут на удивление долго).
Однако для того, чтобы понять, насколько всеобщим является это правило, нужен сравнительный анализ данных по множеству видов. Журнал Nature Communications опубликовал статью китайских биологов, которые провели такой анализ для класса млекопитающих. Млекопитающие — хороший объект для изучения взаимосвязи социальности с долголетием, потому что в пределах этого класса имеется огромная изменчивость по обоим показателям: от одиночек до эусоциальных видов и от долгожителей вроде гренландских китов, которые, возможно, живут до 200 лет, до некоторых видов землероек, не доживающих в идеальных условиях даже до своего третьего дня рожденья.
Авторы собрали литературные данные по социальности и долголетию (максимальной ПЖ) для 974 видов млекопитающих. По уровню социальности зверей разделили на три категории: преимущественно одиночные (497 видов), живущие обычно парами (115 видов) и живущие в основном группами (412 видов). 50 видов были отнесены сразу к двум категориям (например, живущие примерно с одинаковой частотой поодиночке или парами). При отнесении вида к той или иной категории учитывались только взрослые особи (то есть детеныши, живущие вместе с матерью или родителями, в зачет не шли). «Группами» считались объединения трех и более половозрелых особей.
Долголетие в одних расчетах рассматривали как количественный признак, в других — как качественный (бинарный). В последнем случае виды с максимальной ПЖ до 26 лет (включительно) считались короткоживущими, свыше 26 лет — долгоживущими. Эта условная граница соответствует третьему квартилю, то есть 25% видов оказываются «долгоживущими», 75% — «короткоживущими». Если проводить границу в другом месте, например, по медианному значению ПЖ (17 лет), результаты получаются такие же — это авторы проверили. Альтернативный способ анализа ПЖ состоял в том, что делалась поправка на массу тела (потому что крупные звери в среднем живут дольше мелких), и тогда использовалось не абсолютное, а относительное значение максимальной ПЖ. Это тоже не повлияло на результаты.
Филогению рассмотренных видов (топологию эволюционного дерева) взяли из системы TimeTree (см.: S. Kumar et al., 2017. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times). Проверялась и другая версия филогении млекопитающих, что, как можно догадаться, тоже на выводы не повлияло.
Собранные данные в компактном виде представлены на рис. 1. Для обработки таких данных разработаны мощные пакеты программ, где учтены все тонкости биологической статистики, которыми авторы и воспользовались. Для начала проверили, есть ли в данных по социальности и ПЖ «филогенетический сигнал», то есть в какой степени эти признаки «наследуются» в череде происходящих друг от друга видов. Оказалось, что филогенетически сигнал в обоих случаях есть, причем довольно сильный. Это значит, что признаки хорошо «наследуются», и поэтому родственные виды обычно сходны как по социальности, так и по ПЖ. Иначе говоря, эти признаки в ходе эволюции меняются намного медленнее, чем появляются, сменяя друг друга, новые виды. В дальнейших расчетах делались необходимые поправки на этот филогенетический сигнал.
Максимальная ПЖ — как абсолютная, так и относительная (с поправкой на массу тела) — оказалась значимо выше у видов, живущих группами, по сравнению с одиночками (как и ожидалось). Виды, живущие парами, занимают промежуточное положение, но различия между ними и обоими крайними вариантами не дотягивают до порога статистической значимости.
В эволюции млекопитающих многократно происходили переходы из одного состояния в другое: одиночки становились социальными, долгожители — короткожителями и т. д. Авторы показали, что эволюционные переходы между тремя градациями социальности и двумя градациями долголетия происходили с разной частотой (рис. 2).
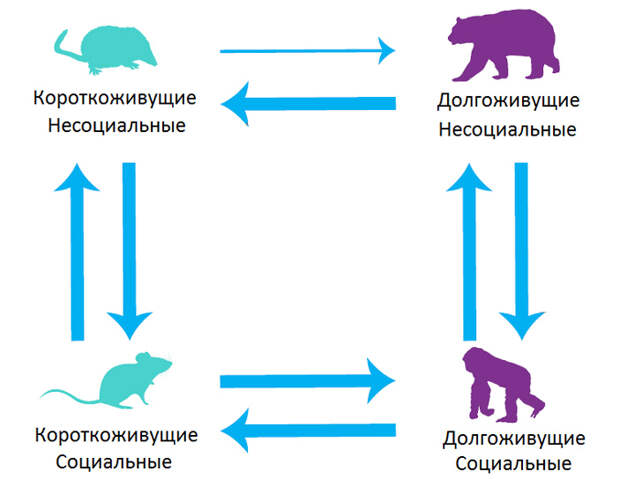
Главный вывод состоит в том, что эволюционные потомки короткоживущих млекопитающих имели гораздо больше шансов стать долгоживущими, если они были социальными. В обратную сторону это не работает: потомки социальных короткоживущих видов становились долгоживущими не чаще, чем потомки несоциальных короткоживущих видов. Это веский аргумент в пользу того, что наблюдаемая у млекопитающих положительная корреляция между социальностью и долголетием объясняется не тем, что долголетие способствует эволюции социальности (такая идея тоже обсуждалась в литературе), а тем, что социальность способствует эволюции долголетия.
Причины такого влияния могут включать не только упомянутую выше защищенность социальных животных от внешних угроз, благодаря чему отбор тщательнее выбраковывает поздно проявляющиеся вредные мутации. Свой вклад могут вносить также факторы, связанные с совместной заботой о потомстве: если старые особи помогают родне выращивать и защищать детенышей, то отбор может поддержать продление жизни даже за пределы репродуктивного периода, вплоть до эволюции настоящей менопаузы (как у людей и некоторых высоко социальных китообразных). Сходным образом могут работать и факторы, связанные с жизненным опытом, социальным обучением и культурным багажом: отбор может поддержать продление жизни, если наличие в группе старых, опытных особей приносит пользу сородичам.
Авторы не остановились на достигнутом (хотя, на мой взгляд, вполне могли бы) и попытались найти «генетический базис» сопряженной эволюции долголетия и социальности у млекопитающих. Вообще говоря, у подобных эволюционных корреляций вовсе не обязательно должен быть какой-то особый генетический базис. То есть не обязательно должны существовать такие гены, эволюционные изменения которых будут приводить к согласованному изменению сразу обоих признаков. Например, долголетие может определяться одними генами, социальность — другими, а сопряженная эволюция объясняется просто тем, что общественный образ жизни способствует отбору на долголетие. Собственно, именно так дело, скорее всего, и обстоит. Но вдруг есть какие-то «волшебные» гены (по аналогии с «волшебными признаками», см. Найден «волшебный признак», способствующий видообразованию у дрозофил, «Элементы», 11.03.2014) или генно-регуляторные сети, изменения которых автоматически дают сразу оба эффекта?
Для поиска таких генов авторы приложили немалые усилия. Во-первых, они изучили транскриптомы мозговой ткани 94 видов млекопитающих, относящихся к 14 отрядам, 39 семействам и 67 родам. Это позволило выявить несколько сотен генов, экспрессия которых в мозге коррелирует с уровнем социальности, и еще несколько сотен генов, экспрессия которых коррелирует с долголетием (например, у короткоживущих активность гена значимо ниже, чем у долгоживущих). Пересечение этих множеств включает 31 ген. Многие из них связаны с иммунной защитой и гормональной регуляцией, что в общем-то не очень удивительно, поскольку иммунная система и гормоны влияют в организме более или менее на всё. Сколько-нибудь убедительных «волшебных» генов, на которые можно было списать сопряженность эволюции долголетия и социальности, среди них не нашлось. Но полученные списки, несомненно, пригодятся в дальнейших исследованиях.
Примерно такие же (не очень внятные) результаты дал и анализ силы отбора, действующего на разные гены у млекопитающих, различающихся по продолжительности жизни и уровню социальности. Отбор оценивали по критерию dN/dS, то есть по соотношению значимых и синонимичных замен в белок-кодирующих участках генов. Это соотношение повышается, если на ген действует положительный (движущий) отбор, поддерживающий значимые замены, оказавшиеся полезными. В итоге получены длинные списки генов, которые подвергаются более сильному (или слабому) отбору у одиночных видов по сравнению с социальными или у короткоживущих по сравнению с долгоживущими. Здесь тоже на пересечении двух множеств убедительных «волшебных» генов не нашлось. Авторы делают вывод, что отбор на социальность и отбор на долголетие могут затрагивать одни и те же регуляторные каскады, метаболические пути и даже одни и те же гены, но молекулярные механизмы эволюционных изменений двух изучаемых признаков все-таки в основном разные. Что ж, это тоже полезно знать.
Главный — и очень важный — результат исследования состоит в том, что, во-первых, подтверждено существование положительной эволюционной корреляции между социальностью и долголетием у млекопитающих. Во-вторых, отвергнута гипотеза о том, что эта корреляция объясняется положительным влиянием долголетия на эволюцию социальности: долгоживущие одиночки становятся социальными не чаще, чем короткоживущие одиночки. В-третьих, подтверждена гипотеза о положительном влиянии социальности на эволюцию долголетия: короткоживущие социальные звери гораздо чаще становятся долгожителями, чем короткоживущие одиночки.
Источник: Pingfen Zhu, Weiqiang Liu, Xiaoxiao Zhang, Meng Li, Gaoming Liu, Yang Yu, Zihao Li, Xuanjing Li, Juan Du, Xiao Wang, Cyril C. Grueter, Ming Li & Xuming Zhou. Correlated evolution of social organization and lifespan in mammals // Nature Communications. 2023. DOI: 10.1038/s41467-023-35869-7.
См. также:
1) Панцири и яды способствуют эволюции долголетия у амфибий и рептилий, «Элементы», 05.07.2022.
2) Быстрое старение у нематоды C. elegans поддерживается родственным отбором, «Элементы», 15.01.2019.
3) Что общего у голых землекопов и «голых обезьян»?, «Элементы», 06.03.2017.
4) Экстракт из старых сородичей ускоряет старение, «Элементы», 20.02.2017.
5) У червя Caenorhabditis elegans старение легко замедлить или ускорить, но трудно изменить его траекторию, «Элементы», 02.02.2016.
6) Как стареют растения, «Элементы», 07.11.2006.
7) Эволюция ускоренного старения как следствие адаптации к неблагоприятным условиям (доклад А. В. Маркова на семинаре «Вопросы эволюции» 27.01.2017).
Свежие комментарии